講演会&セミナー 2014年度
2024年度 |2023年度 |2022年度 |2021年度 |2020年度 |2019年度| 2018年度 | 2017年度 |2016年度 |2015年度 | 2013年度 |2012年度 | 2011年度 | 2010年度 | 2009年度 | 2008年度 | 2007年度 | 2006年度 | 2005年度以前
セミナー記録
2014年12月17日 (水) 16時20分ー17時50分 理学部3号館11番教室 分子生物学科セミナー
演者:斎藤 成也 教授(国立遺伝学研究所 集団遺伝研究部門、総合研究大学院大学 生命科学研究科 遺伝学専攻)
題目: 日本列島の現代人と古代人のゲノムDNA解析
南北に長い日本列島には,北からアイヌ人,ヤマト人,オキナワ人という3種類の人類集団が居住しています.わたしたちは核ゲノム全体に散在している数十万のSNP座位における対立遺伝子情報をもちいて,これら3集団からサンプルした各個体間および世界の他地域の人間の遺伝的近縁関係を推定しました.アイヌ人とヤマト人については,これら2集団が混血をはじめた時期を西暦6世紀と推定し,また集団間で特に大きな対立遺伝子頻度の差があるゲノム領域のなかに,形態的表現型に関与する遺伝子が含まれていることをみいだしました.一方,現代日本列島人にゲノムが引き継がれたと考えられている縄文時代人の核ゲノム塩基配列の一部をはじめて決定し,縄文人が東ユーラシアの中できわめて特異な系統に位置づけられることを発見しました.これらの研究から日本列島人の起源と成立について議論します.
2014年12月9日 (火) 16時20分ー17時50分 理学部2番教室(理学部講義実験棟1階) 分子生物学科セミナー
演者:刀禰 高広 博士(上智大学理工学部)
題目: 微生物遺伝学 ~枯草菌ファージφ29の遺伝子1の機能解析をモデルとして~
様々な生物のゲノム配列が解読され、莫大な量の遺伝情報が蓄積してきている。バイオインフォマティクス的解析を駆使しても、機能不明な遺伝子が多数残されているのが現状である。そうした機能未知の遺伝子の働きを調べる方法として、古典的な遺伝学的手法は未だに有用である。本セミナーでは、サプレッサーや合成致死などの遺伝学的手法を用いて、枯草菌に感染するファージφ29の遺伝子1の機能解析を行った例を紹介する。また、φ29の宿主である枯草菌はグラム陽性菌のモデルとして詳細な研究が行われ、様々な変異体が構築されているが、そうした変異体のうち、ゲノムの様々な非必須領域を欠失させた枯草菌欠失株コレクション(KAO/NAIST株など)を用いた解析から、φ29の遺伝子1と関連がありそうな宿主因子の存在が明らかとなってきた。こうした最新の結果も合わせて紹介する。
2014年11月26日 (水) 16時40分ー18時 理学部3号館11番教室 分子生物学科・環境科学研究センター共催セミナー
演者:Prof. Julian Eaton-Rye(Otago University, New Zealand)
題目: Auxiliary Proteins of Photosystem II ? the Water-Splitting Apparatus
of Oxygenic Photosynthesis
How do these subunits contribute to assembly and function of the mature
Photosystem?
The core of Photosystem II (PS II) is made up of two reaction center proteins, D1 (PsbA) and D2 (PsbD) and two chlorophyll a-binding antenna proteins, CP47 (PsbB) and CP43 (PsbC). These proteins have homologues in anoxygenic photosynthetic bacterial reaction centers; however, PS II has an increased complement of polypeptides. These proteins include ~12 low-molecular-weight (LMW) hydrophobic polypeptides that form a ring around the core and a “cap” of hydrophilic polypeptides that cover the oxygen-evolving center (OEC). These auxiliary PS II proteins were acquired prior to endosymbiotic uptake of cyanobacteria. The crystal structure of cyanobacterial PS II obtained by Nobuo Kamiya and Jin-Ren Shen and co-workers in 2011 has revealed that the cap of the OEC is composed of three polypeptides: PsbO, PsbU and PsbV, and two large loops from CP43 and CP47. In green algae and plants, PsbU and PsbV are absent but two different proteins, PsbP and PsbQ, are present. However, biochemical and genomic studies have now established that there are PsbP and PsbQ homologues in all photosynthetic lineages: and in cyanobacteria the corresponding homologues, CyanoP and CyanoQ, are lipidated at their N-termini. Successful water splitting by PS II carries with it the metabolic cost of light-induced photodamage (chiefly to D1) and the resulting need for a repair cycle to support ongoing O2-evolving activity. It is likely that a major role of the auxiliary proteins is to assist in water splitting and PS II repair. We have investigated the role of the LMW PsbT protein: removal of this subunit increased susceptibility to photodamage and accelerated the rate of incorporation of replacement D1. Moreover, in this mutant, electron transfer between QA and QB, the primary and secondary quinone electron acceptors of PS II, was slowed and the PSII-specific electron acceptor 2,5 dimethyl-p-benzoquinone blocked QA- oxidation. These effects could be prevented, and in some cases reversed, by the addition of HCO3-, a PS II-specific cofactor that binds to the non-heme iron between QA and QB. In addition, we have obtained the X-ray-derived structure of CyanoQ and the NMR structures of CyanoP and a third cyanobacterial lipoprotein, Psb27 (Psb27 is found in all photosynthetic lineages and is involved in the PS II repair cycle). Structural features of these polypeptides will be presented together with new evidence that suggests roles for these proteins at different stages of PS II assembly and in the activity of the holoenzyme assembled in their absence.
解説:
光合成の光化学系II酸素発生複合体は、様々なタンパク質によって保護されています。シアノバクテリアでは、以前からPsbO, PsbV, PsbUの3つの表在性タンパク質が知られていましたが、近年、植物様の
CyanoP, CyanoQやPsb27タンパク質も酸素発生系の保護に関与していることがわかってきました。本講演では、これらのタンパク質の構造と機能に関して最新の知見を紹介していただきます。
2014年10月15日 (水) 14時ー15時 理学部3号館11番教室 分子生物学科・環境科学研究センター共催セミナー
演者:Dr.MEILINA ONG ABDULLAH(Principal Research Officer, Advanced Biotechnology
& Breeding Centre,Malaysian Palm Oil Board)
題目: OIL PALM: PRODUCTIVE, VERSATILE & GREEN
アブラヤシは、その生産性からみて地球上で最も大きなバイオマスの一つであり、経済面からも重要な油脂生産植物です。 アブラヤシは20~30kgもある油脂含量30%の果実房を毎月作りだし、25年間生産し続けます。日本の食品の多くにもパーム油が使われています。その生態、生産性、多用途性から最近のゲノム研究の成果まで講演して頂きます。
参考文献: Nature. 2013 ;500(7462):340-4. The oil palm SHELL gene controls oil yield and encodes a homologue of SEEDSTICK. Nature. 2013 500(7462):335-9. Oil palm genome sequence reveals divergence of interfertile species in Old and New worlds. Nature Commun. 2014 Jun 30;5:4106.The oil palm VIRESCENS gene controls fruit colour and encodes a R2R3-MYB.
2014年9月10日 (水) 16時ー17時 理学部3号館11番教室 分子生物学科・環境科学研究センター共催セミナー
演者: 高木優 教授(理工学研究科戦略研究部門)
題目:転写抑制因子を用いた遺伝子機能解析と応用利用
植物の乾燥、塩害等の環境ストレス耐性強化、生産力向上、バイオマス増産、およびバイオ燃料やバイオプラスチックなどの素材により適合した組成に植物を改良することは、ひいては二酸化炭素資源の有効利用に繋がり、食糧増産、二酸化炭素削減および化石燃料削減に貢献することが期待できる。そのためには、機能に関わる遺伝子因子を見いだし、植物機能を改変することが有効な手段である。植物では、遺伝子発現の制御が主に転写レベルで行われていることから、転写の第一段階を制御する転写因子を利用することが植物機能を改変させる有効な手段であることが判っている。しかし、植物ゲノムには、転写因子を含め機能が重複した遺伝子が数多く存在し、遺伝子破壊や相補RNAでは、表現型が現れず、目的とする転写因子の機能解明が容易ではないという問題があった。そこで我々の研究グループでは、転写因子を強力な転写抑制因子に機能変換し、標的遺伝子の発現を抑制することによって欠損型の表現型を誘導するキ メラリプレッサー遺伝子サイレンシング法(Chimeric Repressor gene Silencing Technology: CRES-T)を開発し、これまで困難であった機能重複した転写因子の機能解明を可能にした。このシステムを用いることによって、これまでの変異体では見られなかった代謝経路を有する植物や、環境ストレスに耐性を持つ植物等が作出できることが判ってきた。この技術を利用して花粉の飛ばない植物や、環境ストレスに耐性を持つ植物や、物質生産に優れたこれまでにない新しい特性をもった植物の開発を行っている。本セミナーでは、CRES-T法の概要とこれらが植物機能の改良に有効なツールであることを紹介する。加えて実用的な植物改良とは何かについて一緒に考えてみたい。(写真はキメラリプレッサーを用いて作成した八重咲きシクラメン:北興化学(株)との共同研究)
メラリプレッサー遺伝子サイレンシング法(Chimeric Repressor gene Silencing Technology: CRES-T)を開発し、これまで困難であった機能重複した転写因子の機能解明を可能にした。このシステムを用いることによって、これまでの変異体では見られなかった代謝経路を有する植物や、環境ストレスに耐性を持つ植物等が作出できることが判ってきた。この技術を利用して花粉の飛ばない植物や、環境ストレスに耐性を持つ植物や、物質生産に優れたこれまでにない新しい特性をもった植物の開発を行っている。本セミナーでは、CRES-T法の概要とこれらが植物機能の改良に有効なツールであることを紹介する。加えて実用的な植物改良とは何かについて一緒に考えてみたい。(写真はキメラリプレッサーを用いて作成した八重咲きシクラメン:北興化学(株)との共同研究)
2014年9月2日 (火) 16時ー17時30分 理学部3号館11番教室 分子生物学科・環境科学研究センター共催セミナー
演者: 上田貴志 准教授(東京大学大学院理学系研究科)
題目: 植物に学ぶ膜交通経路の多様化機構
ゴルジ体やエンドソームなどの単膜系オルガネラは,小胞や小管を介した物質輸送システム,いわゆる“膜交通”を介し,互いに内容物や膜成分の交換をおこなっている(図参照)。この膜交通は,原始真核生物からヒトや植物を含む現存の生物に至るまで保存された,真核細胞に普遍的な生命現象である。一方で,それぞれの生物の多様な体制や生命機能に応じ,膜交通の分子機構や生理機能は進化し,洗練されてきたと考えられる。
最近の比較ゲノム解析の結果は,この膜交通経路の多様性が,RAB GTPaseやSNAREをはじめとする膜交通実行分子の多様化によりもたらされたことを示唆している。しかしながら,RABやSNAREがどのような分子進化を経て新たな機能を獲得し,それらがどのように膜交通網を多様化させたのかについては明らかとなっていない。我々は,植物細胞における膜交通の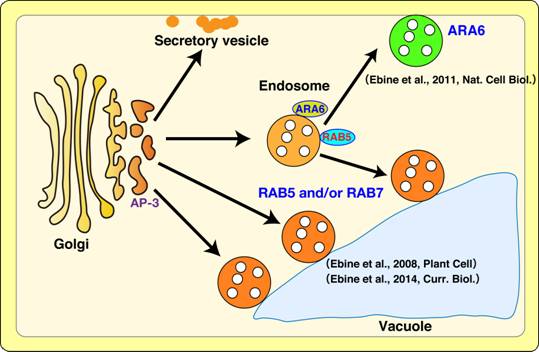 分子メカニズムを明らかにするとともに,その多様化と形質の進化がどのように関連しているのかを解明するべく研究を行っている。 植物における膜交通経路の多様化がどのように起こったのかを知るためには,植物がその進化の過程で独自に獲得した膜交通制御因子の解析が有効である。我々は,植物特異的なRAB
GTPase(ARA6)とSNARE(VAMP727)に注目し,これらの機能を,バイオイメージングをはじめとする様々な手法を用いて解析してきた。本セミナーでは,植物が進化の過程でどのようにポストゴルジ輸送経路を多様化させ,それがどのような植物の高次機能と関連しているのかについて最新の知見を紹介したい。
分子メカニズムを明らかにするとともに,その多様化と形質の進化がどのように関連しているのかを解明するべく研究を行っている。 植物における膜交通経路の多様化がどのように起こったのかを知るためには,植物がその進化の過程で独自に獲得した膜交通制御因子の解析が有効である。我々は,植物特異的なRAB
GTPase(ARA6)とSNARE(VAMP727)に注目し,これらの機能を,バイオイメージングをはじめとする様々な手法を用いて解析してきた。本セミナーでは,植物が進化の過程でどのようにポストゴルジ輸送経路を多様化させ,それがどのような植物の高次機能と関連しているのかについて最新の知見を紹介したい。
2014年4月23日 (水) 16時20分ー17時50分 理学部3号館11番教室 分子生物学科セミナー
演者: Dr. Jenny Mortimer(理化学研究所)
題目: Breaking down walls: unexpected outcomes in the quest for improved biomass
Issues such as climate change, fuel security and diminishing fossil fuel reserves have led to an increased interest in alternative sources of energy. In particular, one important problem is how to replace petroleum products with a renewable liquid transport fuel ("biofuel"). Current biofuels on the market are derived from the fermentation of plant soluble sugars e.g. maize starch or sugar cane sucrose into ethanol. However, both of these plant sugars are also an important global food source, and large-scale utilisation for food production has proved controversial. An alternative approach is to use the sugars locked up in the polysaccharides of the plant cell wall. The cell wall forms the majority of plant biomass, but the components are complex and difficult to break down to their monosaccharide components, and their structure and synthesis poorly understood.
Glycosylation, unlike protein or nucleic acid biosynthesis, has no obvious template, and yet assembly isn't random. Moreover, misglycosylation events can have serious impacts on plant growth and development. My research aims to understand how the flow of substrates for glycosylation is controlled, how they are assembled into polymers, and how these polymers are integrated into the cell wall. Alongside this, I am attempting to integrate this knowledge in order to improve strategies for biomass deconstruction. This has also led me to work on associated biosynthetic pathways, such as the sphingolipid glycosylation, which occurs in the same organelle (the Golgi) and requires the same substrates, but seems to utilise different substrate pools.
Ref: Mortimer et al. (2013) Plant Cell