遺伝情報研究室
カルジオリピンの働き
バクテリア(細菌)の細胞も、私たちの体をつくっている細胞と同じ、りっぱな「細胞」です。DNAからできた染色体が、(核膜に包まれた「核」でなないけれど)細胞の中のしかるべき位置に、しっかりと陣取っているなど、細胞のなかみは秩序だった構造をもっています。その細胞が、きちんと秩序だったやりかたで、細胞分裂をおこなって、増殖する点でも、私たちの体をつくっている細胞と同じです。
細胞のなかみを包んでいる細胞膜は、脂質(リピド)からできています。脂質の間にいろいろなタンパク質が埋め込まれていて、大切な働きをしています。ですから、細胞膜の中のタンパク質も、ちゃんと決まった場所、たとえば、細胞分裂に働くタンパク質は、細胞分裂が起こる細胞中央部に、陣取って働きます。
でも、脂質は、細胞膜をつくっている素材にすぎなくて、単細胞のバクテリアでは、細胞膜全体でどこも区別がないもの、と思われてきました。
ところが、脂質の一種であるカルジオリピンを染める蛍光染色剤(ノニルアクリジンオレンジ)で、枯草菌の細胞を染めてみると、細胞膜が一様に染まるわけではなく、細胞分裂でできる隔壁がつよく染まることがわかりました。
紹介した写真(上)は、ふつうの細胞分裂(細胞を真ん中で2つに分ける)とは違って、胞子をつくりはじめるために、細胞の端っこのほうで、分裂しているようすです。この「不等」分裂でも、隔壁にカルジオリピンがたくさんあることがわかります。写真(下)は、不等分裂のあと、胞子をつくりあげるために、大きいほうの細胞(母細胞)が、小さい方の細胞(胞子になる細胞)を包みこもうとしているところです。この包みこみの過程の細胞膜にもカルジオリピンがたくさん含まれています。カルジオリピンは、胞子をつくるときや、できあがった胞子が発芽するときに、大切な働きをすることが、わかってきています。もう少し詳しく知りたい人は、研究室紹介ページ ![]() もご覧ください。
もご覧ください。
![]()
バクテリアの脂質
寒天培地には、X-Galと略称される物質を入れてあります。この物質は、乳糖分解酵素(β-Galと略称)によって分解されると、青い色を出します。大腸菌は本来β-Galの遺伝子をもっていて、乳糖を利用したいときにだけ働きだしますが、この遺伝子を、人工的にcpsという遺伝子のうしろにつなぎかえました。すると、乳糖を利用したいかどうかとは関係なく、cps遺伝子が働きだす(発現する)ときに、β-Galの遺伝子も働きだし、X-Galを分解できるようになります。写真の4つの菌株は、それぞれいろんな突然変異をもっているのですが、4つとも共通に、そのような人工的な遺伝子のつなぎかえをしてあります。それで、青くなっている2つは、cps遺伝子が発現していることがわかります。
大腸菌は、Rcsシステムという、細胞外の環境の変動によるストレスに応答して遺伝子の発現を制御するための情報伝達経路をもっていて、このシステムがスイッチオンになると、cps遺伝子が発現します。つまり、青くなっているのは、Rcsシステムがスイッチオンになっているからです。(Rcsシステムは、本来は、ほ乳動物の腸から外へ排出されたときに、乾燥・低温・物体表面への接着などに応答して、スイッチオンとなります。)
でも、同じ寒天培地で培養しているのだから、4つの菌株の細胞外環境は、同じ。写真の上半分の2つの菌株でRcsシステムがスイッチオンになっているのは、環境が違うためではなくて、余計にもっている突然変異のためです。
右下の菌株は、β-Galの遺伝子をcpsという遺伝子のうしろにつなぎかえただけ。右上の菌株は、さらに、Rcsシステムの遺伝子に、細胞外環境と関係なくスイッチオンとなりっぱなしになる、突然変異が起きています。
左下の菌株は、β-Galの遺伝子とcps遺伝子とのつなぎかえに加えて、lppという遺伝子に突然変異をもっています。左上の菌株は、さらに、細胞膜を構成する脂質(リピド)のうち「酸性リン脂質」と総称されるものがつくれなくなる突然変異をもっています。(酸性リン脂質がつくれないと、同時にlppもダメになっていなくては生きられないので、lpp遺伝子の変異をもたせてあります。)
左上の菌株では、酸性リン脂質がつくれないために、Rcsシステムがスイッチオンになっています。酸性リン脂質がつくれないと、細胞膜の構造が変わって、それがストレスとなって、スイッチオンになっているのでしょう。こんな実験結果から、Rcsシステムが環境ストレスを感知する仕組みや、細胞膜を構成している脂質の機能を解明する手がかりを見つけようとしています。バクテリアの脂質やその合成酵素についてもう少し詳しく知りたい人は、研究室紹介ページ ![]() もご覧ください。
もご覧ください。
![]()
生体物質研究室
答えは×です。シロイヌナズナは、ナズナや菜の花、ダイコンと同じアブラナ科の植物で、見た目はナズナによく似ています(写真参照)。しかし、ナズナは、種子が入っている果実がハート形の短角果であるのに対して、シロイヌナズナは棒状の長角果です。ナズナは果実の形が三味線のバチに似ていることから、ペンペングサ、とも呼ばれますが、シロイヌナズナの果実は三味線のバチには似ていません。また、ナズナは果実を折り曲げるとカラカラといういい音がしますが、シロイヌナズナではナズナほどいい音は出ません。ですから、シロイヌナズナはペンペングサ(ナズナ)の親戚ですが、ペンペングサではありません。今度、空き地や田圃、畑地の近くを歩くことがあったら、ナズナやシロイヌナズナを探して、果実の形を見比べてみてください。
![]()
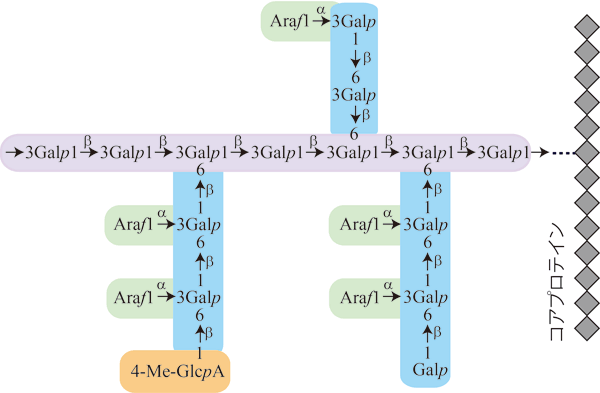
AGP糖鎖の構造
プロテオグリカンというのは、タンパク質に多糖類やオリゴ糖が結合した生体高分子の一種です。私たちヒトの細胞も様々なプロテオグリカンで覆われていますし、植物にもプロテオグリカンが存在します。
プロテオグリカンで面白いのは、その構造と生理的な役割です。植物の細胞膜や細胞壁に普遍的に存在するアラビノガラクタン-プロテイン(以下AGPと呼びます)というプロテオグリカンは、組織や発育段階によって糖鎖部分の構造が変化します。また、AGPは細胞接着や細胞生長、形態形成、細胞死、生殖などに関わることが知られていて、細胞壁の構造多糖類というよりは、情報分子として機能していると考えられます。しかしながら、AGPは生理的な機能に必要な構造(エピトープ)や、その作用機作がまだわかっておらず、謎(?)のプロテオグリカンです。
生体物質研究室では、様々な糖鎖分解酵素やその遺伝子を使ってAGP糖鎖の構造を人為的に改変することで、AGP糖鎖の生理的な役割を調べています。また、各種分析装置を駆使してAGP糖鎖の詳細な構造を調べています。
AGP糖鎖の構造は下図のとおりです。
葉の表面の毛(トライコーム)で発現するAGP(緑色)
![]()
分子統御研究室
答えは、枯草菌の胞子です。透過型電子顕微鏡という、極めて小さなものでも高解像度で観察できる顕微鏡で、胞子を輪切りにして観察しています。原理を簡単に説明すれば、透過型という名の通り、密度の濃い部分は透過率が低くなるので、黒っぽく見えるというわけです。
普段の枯草菌の細胞は大腸菌と同じような長方形をしていますが、その同じ細胞が胞子形成という過程をへて、このような全く形の異なる細胞に変化するのです。層のよう中央の暗い部分にゲノムDNAやタンパク質がコンパクトに納められていて、コアと呼ばれる胞子の心臓部分です。そのコアの外側に、白く抜けている部分がありますが、コルテックスと呼ばれる糖タンパク質でできたかごのようなものです。さらにその外側に灰色の部分と黒い部分の2つの層があります。これらはスポアコートと呼ばれる蛋白質の層で、いわば固い殻のようなものです。このような複雑な階層構造を作って、コアという心臓を守っているのです。
枯草菌は、ある程度の環境変化であれば、遺伝子発現を変化させて、その環境に適応して生育することができます。しかし、生育できないような過酷な環境にさらされた時には、時として胞子という複雑な形態に分化することによって、眠りにつき生きながらえるわけです。私たちの研究室では、胞子形成機構も含め、枯草菌が様々な環境変化に対して、どのようにその変化を感じ取って、どのようにその変化に適応・対応しているのか、といった環境応答の分子機構を遺伝子の発現制御の視点から総合的に解析しています。詳しくは私たちの研究室紹介のホームページ ![]() をご覧ください。
をご覧ください。
![]()
細胞生化学研究室
水のない砂漠で植物が生きられるのは、乾燥した気候に適応した特別な光合成をしているためです。
まず一口に砂漠といっても、全く雨の降らない砂漠はそれほど多くありません。一年のうち限られた期間(雨期)にだけですが、わずかな降雨はあります。また、夜は急激に温度が下がるため相対湿度が上がり、夜露が付いたり霧が立ちこめたりすることさえあります。ですから、「さばく」は漢字で「沙漠」とも書きます。水(さんずい)の少ない土地、という意味です。
そのような乾燥した土地では、植物たちはさまざまな工夫をして生き延びています。例えば、短い雨期に一斉に種子から芽を吹き花まで咲かせてまた種子の形で乾期をしのいだり、細胞の中に保護剤となる特別な物質をためて葉をつけたままカラカラになって乾燥に耐え、水が得られるとまた光合成を始める「復活草(イワヒバ)」と呼ばれるものもいます。
一方、サボテンを初めとする砂漠の植物の多くは、得られる機会の少ない貴重な水を節約できる特別な光合成 -CAM型光合成を行っています。CAM型光合成を行う植物は、昼間、葉の気孔を閉じ乾燥から身を守り、夜に気孔を開け大気中の二酸化炭素を吸収します。しかし、二酸化炭素を糖分に変えるためには光のエネルギーが必要なはずです。サボテンたちはいったいどうやってこの矛盾を解決しているのでしょうか。私たちの研究室では、このCAM型光合成の仕組みを分子のレベルで明らかにしたいと考え、研究しています(Link ![]() )。
)。
![]()

イネ科の草の葉(平行脈ですね)を脱色して細胞壁を染色しました。色の赤いところが細胞壁で、特に維管束部分は細胞壁が厚いので、そこが赤く染まっています。左上がオオムギ、左下がトウモロコシです。肉眼では同じように見える平行脈でも、維管束の数はかなり違い、オオムギでは左右の濃いところと、真ん中の薄赤いところの3本しかありませんが、トウモロコシでは10本以上見えています。維管束の間隔がずいぶん違いますね。間隔が広いのと、狭いのでは光合成のやり方も違います。オオムギはC3光合成、トウモロコシはC4光合成をしています。右側のトダシバは中間のタイプですが、光合成のタイプはどちらでしょうか?実は、4本の維管束の間に縦に並んだ薄赤い細胞が2本ずつ見えていますが、これが維管束に(ある意味で)相当します。C4光合成について詳しくはこちら:(Link1 ![]() ) (Link2
) (Link2 ![]() )
)
![]()
地球上のあらゆる環境に細菌が棲んでいます。地表から地下数キロメートル、高温になって生物が生きられなくなる深度まで、海底ももちろん、さらに生物・人体の表面・体内、あらゆるところに細菌が住み着いています。この写真は生ゴミ堆肥の細菌を調べた結果です。DNAを抽出してきて、リボソームRNAの特定部分をPCRで増幅します。その中にいる細菌種すべてのDNAの混合物が得られました。これを特殊な方法の電気泳動(寒天の中で電流を流して、電荷を持つ分子を分ける方法)で上下方向に分けたところです。濃いバンド、薄いバンドと、堆肥の中にいた細菌の存在量パターンを一目で見ることができます。1が完成した堆肥、2が不良発酵堆肥、3が良好発酵堆肥と、発酵がうまくいっているかどうか、一目で見ることができます。この方法を英語でDGGE (denaturing gradient gel electrophoresis)と言います。
![]()
代謝学研究室
みなさんは、光合成のことはよくご存知ですね。光エネルギーを用いて二酸化炭素から糖を合成する反応のことです。光合成というと、植物の葉にある緑色のクロロフィルが光(エネルギー)を吸収するんだ、とおぼえている人が多いと思います。ところが、シアノバクテリアや紅藻にはクロロフィル以外にも光を吸収するための色素が存在し、そのうちの一つはフィコシアニンとよばれるタンパク質で、写真のように大変美しい青色をしています。私たちは、このタンパク質に魅せられて、もっともっとこのタンパク質と、このタンパク質が多数集まってできるフィコビリソームと呼ばれる巨大な集光装置(下図参照)のことを知りたいと思って、研究をしています。より詳しくは私たちの研究紹介ページ ![]() をご覧下さい。
をご覧下さい。
![]()
環境応答研究室
これは、DNAマイクロアレイという、近年新しく開発された方法です。
写真上で赤、緑、黄色に光っているスポットは、スライドグラス上に、ある生物のDNA断片を貼り付けたものです。たとえば、ゲノムの全塩基配列が解読されている生物なら、1つ1つの遺伝子を1スポットとして貼り付けることができます(全遺伝子を貼りつけると数千スポット以上となる)。このようにして作られたDNAマイクロアレイをどのように使うのか、二つの条件下での遺伝子発現の違いを調べる方法を例にとって、説明します。
- 通常条件下、ストレス条件下で培養した生物からmRNAを取り出す。これら二つの条件下では、遺伝子の働き方が違っていると予想されるが、その違いをこのDNAマイクロアレイ法で見いだそうとしているわけです。
- これらのmRNAから(通常の転写とは逆に)DNAを合成する。このときに蛍光を発する物質を取り込ませることにより、合成されたDNA(cDNAという)に「色」をつける。例えば通常サンプル由来のcDNAは赤色、ストレスサンプル由来のcDNAは緑色の蛍光を発するようにしておく。なお、合成されるcDNAは元の細胞でのmRNAの量を反映する。
- これらのcDNAをDNAマイクロアレイにふりかける。すると、それぞれのcDNAは、そのcDNAの元となった遺伝子(マイクロアレイ上の1スポット)と対を作って結合する。この結合は、二重らせんができるときの塩基対の結合と同じ仕組み。
- 各スポットの蛍光の強さを調べる。あるスポットで赤と緑の蛍光の強さが同じだった場合、その遺伝子には通常、ストレス両サンプルのcDNAが等量結合していること、つまりその遺伝子はストレスの有無によらず働いていることが分かる。また、別のスポットで緑の蛍光のみが検出された場合、その遺伝子に対応するcDNAはストレスサンプルにしか存在しないこと、つまりその遺伝子がストレス条件下で活発に働き出す、ということがわかる。
私たちはこのDNAマイクロアレイ法を用いて、シアノバクテリアがいろいろと異なった環境のもとで、遺伝子の発現をどのように調節しているかを調べています。
![]()
左側の培養管には弱い光、右側の培養管には強い光のもとで育てたシアノバクテリアが入っています。細胞内にあるクロロフィルなどの光合成色素の量が違うため、このように色が違って見えます。
光合成をする生物は、光エネルギーを集めるため、また光合成反応を行うための装置(光合成色素を結合したタンパク質複合体)を持っています。これらの装置の量は、弱光下では、効率よく光を集めて光合成を行うために増加し、強光下では過剰な光によるダメージを避けるため(光合成をする生物だって日焼けするのです)、減少します。それらの調節の結果が、私たちの目には、培養液の色の違いとして見えているわけです。
それではシアノバクテリアは、光の強さをどのように感じ取って、どのように、光合成装置の量を調節しているのでしょうか? 私たちはこの調節の過程を、遺伝子レベルから解明しようと、研究を行っています。もっと詳しい研究内容を知りたい方は、こちら ![]() をご覧下さい。
をご覧下さい。
![]()
植物分子生理研究室
脂質とは生体から有機溶剤で抽出される物質の総称です。(Link ![]() )
)
![]()
膜脂質の「膜」とは、生体膜の「膜」から由来します。つまり、膜脂質とは、細胞膜や細胞内膜構造体(小胞体やミトコンドリア、葉緑体など)に由来する脂質のことです。
![]()
膜脂質の構造を簡単に書くと次のようになります。
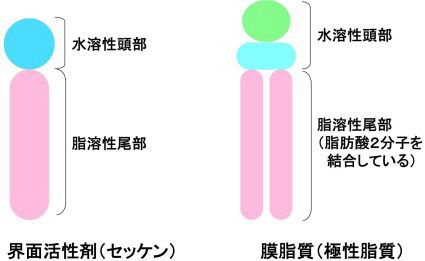 |
膜脂質は、セッケン分子のように水に溶けやすい部分(水色と黄緑色;水溶性頭部)と水に溶けにくい部分(ピンク色;脂溶性尾部)から構成されます。セッケン分子の脂溶性部分は、脂肪族炭化水素鎖ですが、膜脂質の脂溶性部分は、グリセリン骨格(薄い水色)に結合した2本の脂肪酸からできています。 |
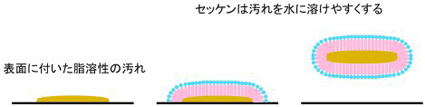 |
セッケンは、脂溶性の油汚れを水に溶けやすくする性質があります。 |
 |
一方、膜脂質はセッケンのようなはたらきを持っていませんが、水溶性部分と脂溶性部分を互いに会合して脂質二重膜(lipid bilayer)と呼ばれる構造体を作ります。脂質二重膜は、生体膜の構造のモデルと考えられています。 |
![]()
答え : 3.10種類
膜脂質は大きく分けて、グリセロ脂質とスフィンゴ脂質に分けられます。スフィンゴ脂質は動物に多い脂質で、分子の種類も多様です。以下には、より基本的で、普遍的な脂質であるグリセロ脂質について、その分子構造の多様性を解説します。
グリセロ脂質には、グリセロリン脂質とグリセロ糖脂質があります。グリセロリン脂質は、6種類(ホスファチジルコリン、PC;ホスファチジルエタノールアミン、PE;ホスファチジルイノシトール、 PI;ホスファチジルグリセロール、PG;ホスファチジルセリン、PS;ホスファチジン酸、PA)知られており、グリセロ糖脂質は、3種類(モノガラクトシルジアシルグリセロール、MGDG;ジガラクトシルジアシルグリセロール、DGDG;スルホキノボシルジアシルグリセロール、 SQDG)が知られています。従って、グリセロ脂質だけで最低9種類の脂質クラスが知られています。
グリセロ脂質のグリセリン骨格には、2つの脂肪酸がエステル結合しています。生体膜の脂肪酸の種類は、主なもので、パルミチン酸(16:0), ステアリン酸(18:0), オレイン酸(18:1), リノール酸(18:2), リノレン酸(18:)3など5種類知られています。ここで、N:Mで表される記号は、炭素数Nと二重結合の数Mからできています。16:0とは、炭素数が16で、二重結合をもたない脂肪酸、すなわち、パルミチン酸のことです。
グリセロ脂質に結合する任意の2つの脂肪酸の組み合わせを考えると5x5=25通り存在します。脂質の種類が9種類ですから、単純に考えると9x25=225種類の脂質分子種が存在することになります。しかし、実際は、グリセリン骨格に結合する脂肪酸の特異性があるので、30種類前後の脂質分子種が存在するようです。
生体膜には、なぜこのようにたくさんの脂質分子種が存在するのでしょうか?われわれの研究室では、異なる分子構造をもった脂質分子種が生合成されるしくみをあきらかにし、そのしくみを遺伝子レベルで壊すことにより(遺伝学的には、変位遺伝子を取ること)、脂質の生理的はたらきをあきらかにしようとしています。
![]()

図 冷蔵庫から出したバター(左)とマーガリン(右)をパンに塗る
左の写真は冷蔵庫から出したばかり、右の写真は室温で15分間放置した後にパンに塗り広げた。15分経ってもバターはうまく塗り広げられない。
マーガリンに含まれる植物性油脂は、不飽和度が高い、すなわちシス二重結合を2-3個含む脂肪酸の含量が高いので、冷蔵庫の温度(2℃)でも完全に固化しません。一方、動物性の油脂であるバターは、飽和脂肪酸の含量が高いので、冷蔵庫の温度ですでに固化します。従って、冷蔵庫から出したばかりのマーガリンはパンにすぐ塗れるのに、バターはパンに塗りにくいということになります。
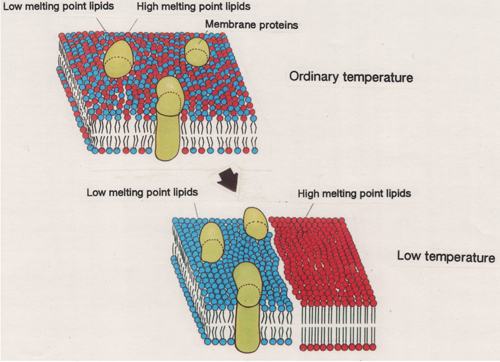
膜脂質には、マーガリンのような脂質とバターのような脂質が存在します。マーガリンのような膜脂質とは、シス二重結合を最低ひとつ含む脂肪酸を最低1分子結合しています(下図では青い色の分子;低融点脂質分子種;low melting point lipids)。一方、バターのような膜脂質とは、飽和脂肪酸を2分子結合しており、低温で固まりやすい性質を示します(下図では赤い色の分子;高融点脂質分子種;high melting point lipids)。低温で膜が固くなると、膜タンパク質(membrane protein)は、柔らかい膜の部分へと排除され、生体膜は正常に機能できなくなると考えられます。植物のように低温環境から逃げることができない生き物では、バターのような膜脂質の割合が高いと、低温で膜の機能に障害が発生し、低温環境を生き延びることが難しくなります。